
VOLUMEN: VI NÚMERO: 14-15
VOLUMEN: VI NÚMERO:
14-15
EMOCIÓN Y MODULACIÓN DE REFLEJOS: NUEVAS PERSPECTIVAS PSICOFISIOLÓGICAS[1][2]
Jaime Vila y María del Carmen Fernández-Santaella
Universidad de Granada
1. El punto de partida: La polémica James versus Cannon
El estudio experimental de la emoción tiene su punto de partida en el artículo que William James publicó en la revista Mind en el año 1884 con el título “¿Qué es una emoción?”. Como en otros muchos aspectos de la psicología, James no sólo fue pionero en el tema de la emoción, sino que también estableció las coordenadas en las que se ha movido la investigación experimental hasta nuestros días. James planteó el tema en términos que hoy llamaríamos neurocientíficos –el de la localización cerebral de las emociones en centros independientes o en centros comunes con otros procesos sensoriales y perceptivos- pero defendió su arriesgada apuesta –a favor de la segunda alternativa- utilizando argumentos puramente psicológicos. Sin embargo, en el momento de formalizar su planteamiento teórico sobre la naturaleza de la emoción, William James quiso apartarse del sentido común provocando en el lector una reacción inmediata de incredulidad. “Nuestra manera natural de pensar sobre las emociones es que la percepción mental de algún hecho provoca la disposición mental llamada emoción y que este estado mental da lugar a la expresión corporal. Mi tesis, por el contrario, es que los cambios corporales siguen directamente a la percepción del hecho desencadenante y que nuestra sensación de esos cambios según se van produciendo es la emoción. El sentido común nos dice que nos arruinamos, estamos tristes y lloramos; que nos topamos con un oso, nos asustamos y corremos; que un rival nos ofende, nos enfadamos y le golpeamos. La hipótesis defendida aquí afirma que este orden de la secuencia es incorrecto, que un estado mental no es inducido inmediatamente por el otro, que las manifestaciones corporales deben interponerse previamente entre ambos y que una exposición más racional es que nos sentimos tristes porque lloramos, enfadados porque golpeamos, asustados porque temblamos, y no que lloramos, golpeamos o temblamos porque, según el caso, estemos tristes, enfadados o asustados. Si los estados corporales no siguieran a la percepción, esta última poseería una conformación totalmente cognitiva, pálida, incolora, carente de calor emocional. Entonces podríamos ver el oso y juzgar que lo mejor es correr, recibir la ofensa y considerar que lo correcto es golpear, pero no podríamos sentirnos realmente asustados o iracundos” (pag. 190, traducción de Elena Gaviria, 1985).
La propuesta de James implicaba que la investigación experimental de las emociones debía asentarse sobre tres pilares: la fisiología, la especificidad y el periferalismo. En primer lugar, la emoción, entendida como la cualidad de la experiencia interna, debe estudiarse registrando y analizando las respuestas fisiológicas. En segundo lugar, tales respuestas conforman patrones corporales específicos para cada emoción. Cada emoción representa, por tanto, una categoría emocional diferente, con su propio patrón de respuestas. En tercer lugar, la fisiología que en última instancia determina las emociones es la fisiología del sistema nervioso periférico, no la fisiología del sistema nervioso central.
Cannon, que siendo estudiante de James en la universidad de Harvard ya había planteado privadamente objeciones a su teoría, debatió públicamente años más tarde los puntos débiles de su planteamiento con sus apasionados seguidores (ver Newman, Perkins y Wheeler, 1930; Cannon, 1931). Cannon fue desmontando prácticamente todos los supuestos de la teoría de James con datos neurofisiológicos de tipo clínico y experimental. El mismo James había indicado que la principal prueba de la veracidad de su hipótesis se encontraría en estudios con pacientes que tuvieran dañadas las vías motoras y sensoriales periféricas, como ocurre en el caso de las personas con lesión medular. Cannon cita el artículo de Dana (1921) en el que una paciente con lesión completa de la médula espinal a la altura de la tercera y cuarta vértebra cervical no sufrió ningún cambio aparente en su capacidad emocional. Cannon añade argumentos basados en datos experimentales sobre la inespecificidad de los cambios orgánicos (los mismos cambios viscerales se producen en estados emocionales muy diferentes), la relativa insensibilidad de las vísceras (las vías interoceptivas aferentes proporcionan un feedback muy pobre), la lentitud de los cambios viscerales en comparación con la rapidez de las reacciones emocionales y, sobre todo, la permanencia de los comportamientos emocionales en animales que han sido sometidos a vagotomías y simpatectomías, esto es, a la separación del sistema nervioso periférico (autónomo) y el sistema nervioso central. Eran críticas aparentemente demoledoras porque los argumentos de Cannon apuntaban a la ignorancia por parte de los defensores de la teoría de James de los fundamentos neurocientíficos de su teoría.
El planteamiento de Cannon mantenía el carácter fisiológico de la teoría de James pero los tres pilares del nuevo modelo eran totalmente contrarios: centralismo (frente a periferalismo), cognición (frente a fisiología corporal) y dimensionalidad (frente a especificidad). En primer lugar, la clave de la emoción estaría exclusivamente en el cerebro, en las interacciones entre el tálamo y la corteza. En segundo lugar, la cualidad emocional sería un proceso básicamente cognitivo. Sentimos miedo no porque corremos sino porque sabemos que la situación es de peligro, aunque al mismo tiempo salgamos huyendo. Por último, las respuestas corporales periféricas sólo contribuyen a dar energía e intensidad a la conducta, pudiendo ser idénticas en emociones muy diferentes. Su posible aportación se limita a una dimensión general de activación, sin conformar patrones específicos de respuesta.
La investigación experimental sobre las emociones, posterior a Cannon, se ha movido entre los tres posicionamientos dicotómicos implícitos en la polémica James-Cannon (periferalismo-centralismo, especificidad-dimensionalidad y fisiología-cognición), poniendo mayor o menor énfasis en un extremo o en el otro en función del paradigma dominante (conductista o cognitivo), pero sin que los datos terminaran de dar la razón a ninguno de ellos. Un ejemplo ilustrativo son los estudios con lesionados medulares. Años después de publicar Cannon sus críticas citando el artículo de Dana de 1921, un investigador, él mismo tetrapléjico, realizó un estudio con 25 lesionados medulares divididos en grupos en función del nivel de su lesión (Hohmann, 1966). Los participantes en la investigación evaluaron retrospectivamente la intensidad de cuatro emociones (miedo, ira, tristeza y sentimentalismo) antes y después de la lesión. Hohmann encontró que la intensidad de las emociones, todas menos sentimentalismo, había disminuido significativamente después de la lesión, siendo las disminuciones más pronunciadas en los grupos con lesiones medulares más altas. Estos datos volvían a dar la razón a James, bien en la versión dura de su teoría (sin feedback no hay emoción) bien en su versión blanda (la disminución del feedback periférico disminuye la intensidad de la emoción). Sin embargo, y pese a que el estudio de Hohmann se recoge en casi todos los textos de psicología general como dato concluyente, varios estudios posteriores han sido incapaces de replicar sus resultados. Al contrario, los últimos datos apuntan que en personas con lesión medular, comparadas con personas sin lesión medular, la intensidad de las emociones no sólo no disminuye, sino que aumenta (Cobos, Sánchez, García, Vera y Vila, 2002). Planteada en los términos dicotómicos periferalismo frente a centralismo –y lo mismo ha ocurrido con el debate especificidad/dimensionalidad y fisiología/cognición- la pelota del partido James versus Cannon sigue todavía en juego.
2. La emoción durante el predominio conductista y cognitivo
El conductismo se sintió incómodo con el tema de la emoción. Por una parte, su rechazo del introspeccionismo le hacía dudar de que fuera correcto el punto de partida tanto de James como de Cannon: que el objeto de la psicología de la emoción fuera explicar la cualidad de la experiencia consciente, esto es, la fenomenología de la emoción. Por otra parte, entendía que toda conducta es conducta motivada apetitiva o aversivamente y, por tanto, que los fenómenos emocionales asociados a la cualidad positiva o negativa de los estímulos podían ser importantes en la explicación de la conducta. El conductismo de Watson y de la escuela de Hull, por ejemplo, dirigieron el estudio de la conducta hacia temas típicamente emocionales: el miedo, las fobias, la frustración, el castigo. Pero su estudio se enfocó desde un planteamiento que podríamos llamar híbrido entre las posturas de James y de Cannon. La identificación de la emoción con las respuestas fisiológicas periféricas (James) permitía deshacerse de la introspección como metodología de investigación. Los registros fisiológicos eran suficientes para obtener la medida objetiva de la emoción. Por otra parte, la identificación de las respuestas fisiológicas periféricas con la dimensión de intensidad o energía de la conducta (Cannon) permitía al conductismo de Hull conectar el estudio de la emoción con el estudio del impulso (drive), entendido como fuerza motivacional, y defender la existencia de un impulso general al que contribuían tanto los estados emocionales (por ejemplo, el miedo) como los estados motivacionales (por ejemplo, el hambre). El planteamiento de Cannon era, además, atractivo para el conductismo puesto que se fundamentaba en la investigación animal, donde la introspección no tiene cabida. Para el conductismo, la emoción terminó siendo el estudio de la conducta motivada apetitiva o aversivamente. Emoción era igual a motivación y motivación igual a activación fisiológica. Por consiguiente, no era imprescindible que la psicología científica siguiera manteniendo el dudoso y ambiguo concepto de emoción.
La psicología cognitiva, posterior al conductismo, también se sintió incómoda con los temas emocionales. Para la psicología cognitiva del procesamiento de la información, la emoción es básicamente un elemento perturbador; algo que no contribuye al funcionamiento eficaz del sistema. De ahí que se haya ignorado su existencia o se haya intentado reducir a otro proceso cognitivo. Los procedimientos de investigación de los temas cognitivos son procedimientos carentes de emoción. Se pueden repetir una y otra vez, bajo el supuesto de que el estado afectivo de la persona a lo largo de las tareas es invariablemente neutro, algo que se intenta garantizar seleccionando estímulos emocionalmente fríos. Hubo, sin embargo, algunas excepciones. Una de las más interesantes fue la que surgió en torno a los estudios sobre memoria dependiente del estado afectivo, esto es, sobre cómo el estado emocional de la persona en el momento del aprendizaje y en el momento del recuerdo facilita o dificulta la memoria (Bower, 1981).
En el modelo de Bower, la emoción se incorpora al modelo cognitivo como un nodo o código representacional. El estado emocional en que se encuentra la persona durante el aprendizaje se almacena en la memoria, junto con los otros nodos de información que se están aprendiendo, justo en el mismo momento de la codificación de la información. Cuando posteriormente se activa esa emoción, automáticamente se activa el resto de nodos de la red produciéndose el recuerdo del material almacenado junto con la emoción. De ahí que el recuerdo de determinados sucesos ocurra más fácilmente cuando nos encontramos en el mismo estado emocional que teníamos cuando se produjeron tales sucesos que cuando nos encontramos en un estado emocional totalmente distinto. La teoría bio-informacional de la emoción de Lang (1979) se basa en el modelo de Bower. Lang amplió la idea de nodo emocional a una red de nodos y proposiciones emocionales. Además, caracterizó la red emocional por su necesaria inclusión de proposiciones de respuesta (entre ellas, las respuestas fisiológicas periféricas), conectando la red cognitiva con los circuitos cerebrales activadores de los componentes somáticos y viscerales de la emoción. En este sentido, el modelo de Lang se apartó del centralismo puro de los psicólogos cognitivos integrando lo periférico y lo central en un marco conceptual que superaba la vieja dicotomía periferalismo-centralismo.
Los planteamientos de Bower y Lang constituyen excepciones dentro de la psicología cognitiva del procesamiento de la información que, durante décadas, siguió desinteresándose por los temas emocionales. Al desinteresarse por la emoción, la psicología cognitiva cometió el mismo error que el conductismo: centrarse en un aspecto de la conducta, el que tiene que ver con el conocimiento, el pensamiento, la mente cognitiva, dejando las emociones y los sentimientos fuera. Las mismas conductas externas, las acciones, quedaron muy degradadas dentro del paradigma cognitivo. El error fue olvidarse del cuerpo biológico que sustenta la conducta cognitiva asumiendo una postura funcionalista extrema según la cual las funciones inteligentes realizadas por diferentes aparatos u organismos vivos reflejan idénticos procesos subyacentes, no importando el soporte físico del que estén hechos unos y otros. De ahí que para la psicología del procesamiento de la información la metáfora del comportamiento humano sea el ordenador electrónico, no el ordenador biológico.
3. La nueva perspectiva psicofisiológica
Estudiar la actividad del organismo humano desde la perspectiva del ordenador biológico significa investigar lo que hacemos, pensamos y sentimos teniendo en cuenta simultáneamente los mecanismos psicológicos y fisiológicos que sustentan dicha actividad. Tradicionalmente, la psicofisiología ha sido la disciplina comprometida con el estudio de tales relaciones. Sin embargo, la perspectiva psicobiológica o neurocientífica, tal como se ha entendido hasta hace relativamente pocos años, no consiguió entusiasmar a la mayor parte de los psicólogos experimentales. Una razón fue el riesgo de reduccionismo biologicista implícito en su planteamiento. Estudiar la conducta exclusivamente en términos de hardware del sistema era reducir la explicación a funcionamiento neuronal o bioquímico, perdiendo el nivel de análisis y explicación propiamente psicológicos. Por otra parte, las investigaciones se realizaban mayoritariamente con animales –fundamentalmente, moluscos y roedores- cuya analogía con el comportamiento humano, a pesar de sus similitudes en el funcionamiento neuronal y bioquímico, deja bastante que desear.
Una parte de la investigación neurocientífica, sin embargo, sí se realizaba con sujetos humanos, la que terminó denominándose psicofisiológica en sentido estricto. Su objeto de estudio no era conocer el hardware del sistema sino su software. Esta perspectiva se caracterizó por utilizar técnicas de registro fisiológico que no alteraran el funcionamiento psicológico de las personas mientras estas realizaban las tareas experimentales, técnicas de registro conocidas como no invasivas. Son técnicas que captan la actividad de los músculos, las vísceras y el cerebro desde el exterior del cuerpo a través de sensores colocados sobre la piel o próximos a ella. Su finalidad es conocer los mecanismos psicológicos que controlan la conducta, bajo el supuesto de que los procesos psicológicos -percepción, atención, memoria, pensamiento, motivación, emoción, etc.- se reflejan en los cambios fisiológicos periféricos –los precursores logísticos e instrumentales de la conducta-, además de reflejarse en los propios cambios fisiológicos cerebrales. Esta estrategia de investigación se ha ido abriendo paso desde la época de James y Cannon, con no pocas dificultades, perfilándose en los últimos años como la mejor estrategia experimental para estudiar la conducta con referencia al cuerpo biológico, sin caer en reduccionismos biologicistas o funcionalistas.
El creciente éxito de la perspectiva psicofisiológica se ha debido a los impresionantes avances en las técnicas no invasivas de registro fisiológico ocurridos en las últimas décadas. La ciencia, en general, avanza cuando aparecen nuevas tecnologías que permiten estudiar los mismos problemas desde nuevas perspectivas o con nuevos métodos. Las nuevas tecnologías que están revolucionando el campo psicológico son como auténticas ventanas sobre el cuerpo viviente que permiten ver y registrar lo que ocurre en los músculos, el corazón y el cerebro cuando actuamos, sentimos y pensamos. Las más recientes son las que proporcionan imágenes del funcionamiento del cerebro: las técnicas de neuroimágenes. Existen dos tipos generales de técnicas, las basadas en la actividad electromagnética del cerebro y las basadas en su actividad metabólica. Ambas han sido posibles gracias a los avances en el campo de la ingeniería, la electrónica y la informática. Las primeras tienen su punto de arranque en los registros electroencefalográficos y en la obtención, a partir de ellos, de los potenciales corticales evocados. Las segundas tienen su punto de arranque en las técnicas radiográficas y en el registro del flujo sanguíneo cerebral. Se asume que la actividad eléctrica neuronal requiere consumo de energía y que la diferente distribución de la sangre en el cerebro es un fiel reflejo de la actividad neuronal.
Estas técnicas han despertado el interés de muchos psicólogos cognitivos que han descubierto en ellas la posibilidad de indagar y desentrañar directamente las estructuras y operaciones mentales inferidas a partir de las medidas comportamentales clásicas: el tiempo de reacción y el número de aciertos y errores durante las tareas. La llamada neurociencia cognitiva es el resultado de la incorporación por parte de la psicología experimental del procesamiento de la información de las nuevas técnicas psicofisiológicas de registro de la actividad cerebral. Como ha señalado Posner (1999), por primera vez en la historia humana, somos capaces de ver el lugar en el cerebro donde se activan las neuronas cuando las personas normales piensan. Los nuevos instrumentos nos permiten observar la anatomía del cerebro mientras se aprenden y se llevan a cabo las complejas tareas de que son capaces los seres humanos. Pero las nuevas técnicas psicofisiológicas no sólo han despertado el interés de la psicología experimental cognitiva. Prácticamente todas las áreas de la psicología se están interesando por estas nuevas técnicas de investigación que, en el ámbito de la conducta, cumplen el mismo papel que el telescopio o el microscopio cumplieron en el ámbito de la astronomía y la biología: permiten penetrar en las interioridades de la conducta no siempre disponibles a la observación externa o a la auto-observación.
Esta nueva perspectiva se está configurando como una aproximación integradora, el lugar de encuentro de las múltiples disciplinas y subdisciplinas en las que se ha ido dividiendo la psicología desde sus inicios. Psicobiólogos, neuropsicólogos, psicólogos experimentales, psicólogos evolutivos, psicólogos clínicos y psicólogos sociales se están interesando por estas técnicas que, además, requieren de la colaboración de otros muchos profesionales: físicos, ingenieros, informáticos, biólogos, neurólogos, cardiólogos, psiquiatras. Es, sin duda, una nueva forma de hacer ciencia que permite el acercamiento entre disciplinas aparentemente alejadas. En el ámbito específico de la psicología experimental, no sólo está permitiendo introducir de pleno derecho la emoción en lo que tradicionalmente ha sido el dominio casi exclusivo de lo conductual y lo cognitivo, sino que gracias a la incorporación de la emoción se está empezando a asumir de forma explícita el importante papel que el cuerpo biológico juega en la explicación de la conducta. Cada vez es más evidente que no sólo el cerebro cognitivo es importante. También lo es el cerebro emocional, como también lo es el resto del cuerpo: el corazón y los músculos. Además, gracias a la nueva perspectiva psicofisiológica, se están empezando a superar las viejas dicotomías periferalismo-centralismo, especificidad-dimensionalidad y fisiología-cognición que han caracterizado el estudio científico de la emoción desde la época de James y Cannon.
4. Un nuevo marco teórico y metodológico
El marco teórico y metodológico que presentamos a continuación representa un ejemplo de la nueva forma de abordar el estudio experimental de la emoción desde la perspectiva psicofisiológica. En él convergen muchas de las propuestas que se han ido formulando, desde áreas y metodologías de investigación diferentes, sobre los mecanismos de la regulación emocional. Además, constituye el pórtico de los trabajos de investigación que describimos en el apartado siguiente bajo el título entre el corazón y el cerebro.
4.1 El modelo de P. J. Lang
Lang, al igual que otros investigadores, entiende las emociones como disposiciones para la acción que se han ido desarrollando a lo largo de la evolución a partir de reacciones de carácter adaptativo ante situaciones relevantes para la supervivencia (búsqueda de alimento, aproximación sexual, lucha, huida). Por ejemplo, la ira dispondría al ataque, el miedo a escapar, el placer y la alegría a aproximarse. Los mecanismos neurales de estas reacciones se encuentran en el cerebro humano en centros sub-corticales y corticales profundos estando directamente relacionados con dos sistemas motivacionales primarios: el apetitivo y el aversivo. La emoción se produciría cuando se activa cualquier estructura de información en el cerebro que conecte con estos sistemas motivacionales. Es entonces cuando se observarían las manifestaciones de la emoción en el triple sistema de respuesta: el conductual, el expresivo-lingüístico y el fisiológico. El sistema conductual incluye tanto las acciones externas típicamente emocionales (aproximación, ataque, huida) como los efectos de la emoción en la ejecución de tareas no emocionales (atención, memoria, aprendizaje) facilitándolas o dificultándolas. El sistema expresivo-lingüístico incluye tanto la comunicación verbal directa de tipo emocional (expresiones de alegría, tristeza, miedo, ira) como los informes evaluativos sobre los propios sentimientos y afectos. Por último, el sistema fisiológico incluye tanto las respuestas viscerales y somáticas, las cuales constituyen el apoyo logístico e instrumental de las acciones externas, como los propios cambios fisiológicos cerebrales. En el modelo de Lang, estos tres sistemas de respuesta son relativamente independientes. Por ello no se pueden tomar cada uno de forma aislada. Hacerlo así proporcionaría una visión imperfecta y parcial del complejo fenómeno emocional, dado que las correlaciones entre e intra sistemas son, en general, bajas.
El modelo de Lang, además, presta especial atención a la organización estructural de las reacciones emocionales, a sus fundamentos neurofisiológicos y a los mecanismos de activación de la emoción (Lang, 1995; Lang, Davis y Öhman, 2002). Lang asume una organización jerárquica de las reacciones emocionales en la que tienen cabida tanto los aspectos específicos como los aspectos dimensionales. En el nivel inferior de la jerarquía predominan los patrones específicos de la respuesta emocional. Las emociones siempre se manifiestan como patrones específicos de acción dependientes del contexto, por tanto, con una topografía fisiológica y conductual específica. El miedo ante un examen se manifiesta de forma diferente al miedo a que te roben cuando pasas por una calle solitaria. Cada reacción emocional es diferente dependiendo del contexto en el que ocurre. En el nivel intermedio predominan los programas emocionales, integrados por subrutinas de ataque, huida, búsqueda de alimentos, aproximación sexual. Estos programas muestran ciertas similitudes y estereotipias de respuesta ante diferentes contextos dando lugar a las llamadas categorías emocionales: el miedo, la ira o la tristeza. Por último, en el nivel superior predominan las dimensiones emocionales. Todas las reacciones emocionales comparten determinadas características: direccionalidad (tendencia a la aproximación o a la evitación), intensidad (mayor o menor requerimiento de energía) y control (continuidad o interrupción en la secuencia conductual). Estas tres características constituyen las tres grandes dimensiones que organizan el mundo emocional al más alto nivel: valencia (agradable-desagradable), arousal (activado-calmado) y dominancia (controlador-controlado).
En cuanto a sus fundamentos neurofisiológicos, el modelo resalta las bases neurales de la dimensión de valencia (agradable-desagradable) relacionadas con los dos sistemas motivacionales primarios: el apetitivo, dirigido a la conducta consumatoria, sexual o de crianza, y el defensivo, dirigido a la conducta de protección, escape o evitación. Este sustrato neurofisiológico es el que justifica el carácter bipolar de la dimensión de valencia y su primacía sobre las otras dos dimensiones. El circuito neural más estudiado y conocido hasta la fecha es, sin duda, el defensivo siendo una de sus principales estructuras subcorticales el núcleo central de la amígdala (Davis, 1997; Lang, Davis y Öhmna, 2002). Por su parte, la dimensión de arousal no tendría un sustrato neurofisiológico separado, sino que representa bien activación de cualquiera de los dos sistemas –el apetitivo o el defensivo- bien co-activación de ambos sistemas. Por tanto, en cuanto a la dimensión de arousal, los dos sistemas motivacionales primarios podrían funcionar de forma aditiva. Sin embargo, en cuanto a sistemas de acción, el apetitivo y el defensivo funcionarían de forma recíprocamente inhibitoria. La dirección general de la conducta (aproximación-evitación) dependerá de las fuerzas relativas de activación de ambos sistemas. El sistema motivacional dominante –apetitivo o defensivo- será el que termine modulando toda la conducta, ya sean reflejos simples o procesos cognitivos complejos. Por último, la dimensión de dominancia es la que menor peso tiene en el modelo teórico de Lang. En los estudios factoriales es también el factor que menor varianza explica, encontrándose sistemáticamente una correlación positiva entre valencia y dominancia: a mayor valencia positiva mayor control y dominio. No obstante, dado que se trata de un factor independiente que aparece en todos los estudios, su aportación diferencial a la organización emocional y, en definitiva, a la conducta debería seguir siendo objeto de interés por parte de los investigadores.
El modelo presta también especial atención a los mecanismos de activación de la emoción. En humanos, la activación de los circuitos neurofisiológicos de la emoción no sólo se produce por la presencia de estímulos externos relevantes para la supervivencia. Debido a las múltiples conexiones de las estructuras motivacionales primarias (subcorticales y corticales profundas) con las estructuras neurales más recientes (corticales), los circuitos neurofisiológicos de la emoción pueden también activarse por el procesamiento interno de estímulos simbólicos o mediante la activación de memorias afectivas. Este aspecto del modelo de Lang es el que queda recogido en su teoría bio-informacional, a la que nos hemos referido anteriormente. La característica definitoria de una red de información emocional es la presencia de información de respuesta, la que permite conectar la estructura cognitiva con los circuitos neurofisiológicos apetitivos y defensivos. Sólo cuando se activan estos circuitos es cuando el procesamiento cognitivo de la red se puede considerar emocional.
El planteamiento teórico de Lang tiene como especial característica el romper con las viejas dicotomías periferalismo-centralismo, especificidad-dimensionalidad y fisiología-cognición. La organización jerárquica de la emoción y sus mecanismos de activación incluyen elementos que son simultáneamente específicos (nivel inferior) y dimensionales (nivel superior), centrales (circuitos cerebrales) y periféricos (respuestas viscerales y somáticas), y cognitivos (red de información) y fisiológicos (funcionamiento corporal).
4.2 La visualización de imágenes afectivas
La principal fuente de datos a favor de este tipo de modelo procede de investigaciones dirigidas a conocer los circuitos psicológicos y fisiológicos de los sistemas apetitivo y defensivo utilizando como principal paradigma de investigación la visualización de imágenes afectivas. En la vida real, las reacciones emocionales pueden ser provocadas por una gran variedad de estímulos. En contextos de laboratorio, sin embargo, el control experimental y los principios éticos restringen considerablemente el abanico de estímulos que pueden ser utilizados con un mínimo de rigor metodológico y respeto a los participantes en las investigaciones. La identificación de estímulos que difieran cuantitativamente en sus características afectivas y que puedan ser manipulados con facilidad por parte del experimentador, y sin rechazo por parte de los participantes, es un objetivo prioritario en la investigación de laboratorio sobre las emociones. La visualización de fotografías de contenido emocional ha sido uno de los procedimientos más utilizados en la investigación humana precisamente por cubrir parte de estos requisitos. Sin embargo, la visualización de imágenes para inducir emociones tiene también importantes limitaciones metodológicas.
Las imágenes y fotografías afectivas difieren considerablemente de los estímulos que se utilizan en muchas investigaciones con animales para provocar reacciones emocionales primarias: ruidos y calambres. Estos estímulos físicos, cuya intensidad puede ser controlada fácilmente, actúan como estímulos incondicionados que provocan reacciones reflejas: los reflejos de orientación, sobresalto y defensa. Sin embargo, las imágenes y fotografías afectivas tienen la característica de ser estímulos perceptuales complejos, altamente simbólicos, y cuya codificación emocional requiere aprendizaje y elaboración cognitiva. Tienen la ventaja de cubrir una gama más amplia de estados afectivos que los típicos ruidos y calambres —limitados al polo aversivo del espectro afectivo— y de representar características importantes de los valores culturales y sociales transmitidos por los medios de comunicación —arte, cine, televisión, revistas. Este tipo de estímulos visuales recoge muchos aspectos de la vida real —deporte, moda, paisajes, desastres naturales, erotismo, violencia, etc.—, actuando como potentes generadores de emociones. Tienen, además, la ventaja de ser estímulos que pueden ser fácilmente presentados en contextos de laboratorio, tanto en formato analógico como digitalizado, permitiendo el control preciso de su momento y tiempo de exposición.
Sin embargo, tienen el inconveniente de ser estímulos de difícil cuantificación afectiva. La principal dificultad de utilizar fotografías de la vida real para generar afectos es su calibración psicométrica en dimensiones análogas a la métrica física. Además, este tipo de estímulos, debido a su fuerte componente de aprendizaje, tiene la dificultad añadida de variar de acuerdo con el contexto cultural y social de la población a la que se va a aplicar. Los métodos psicofísicos clásicos y modernos se centran en sensaciones cuya dimensión física es fácil de manipular: temperatura, peso, brillo, tono, etc. La métrica de las sensaciones afectivas es ciertamente más difícil. No obstante, existe también una larga tradición psicométrica en áreas afines, tan complejas o más que las emociones, como son las actitudes sociales (Thurstone, 1931), el lenguaje (Osgood, Suci y Tannenbaum, 1957) o la personalidad (Eysenck, 1960). La aplicación de esta metodología a la evaluación de imágenes afectivas es el objetivo del Sistema Internacional de Imágenes Afectivas o IAPS (International Affective Picture System).
El Sistema Internacional de Imágenes Afectivas es un instrumento elaborado inicialmente en el Centro para el Estudio de la Emoción y la Atención que dirige el profesor Lang en la Universidad de Florida y que ha sido adaptado recientemente a la población española en las Universidades de Granada y Castellón (Moltó y col., 1999; Vila y col., 2001). Actualmente incluye 832 fotografías en color, en formato de diapositiva o en formato digitalizado, pertenecientes a diversas categorías semánticas: animales, escenas de la naturaleza, objetos caseros, desnudos, parejas eróticas, caras humanas, cuerpos mutilados, armas, comida, deportes, etc. La construcción del IAPS ha estado guiada por el marco conceptual que hemos resumido en el apartado anterior y por la aplicación de un método psicométrico de construcción de escalas: el Maniquí de Auto-Evaluación o SAM (Self-Assessment Manikin). El SAM utiliza escalas pictográficas –no verbales- de fácil y rápida aplicación (ver figura 1). Proporciona información para cada imagen de las tres dimensiones generales: valencia, arousal y dominancia. Cada escala está formada por una secuencia de cinco figuras humanoides, graduadas en intensidad, representando diferentes niveles de agrado, excitación o dominio, desde un extremo (izquierda) de máximo agrado, excitación o sentirse dominado, hasta el otro extremo (derecha) de máximo desagrado, relajación o sentirse dominador.
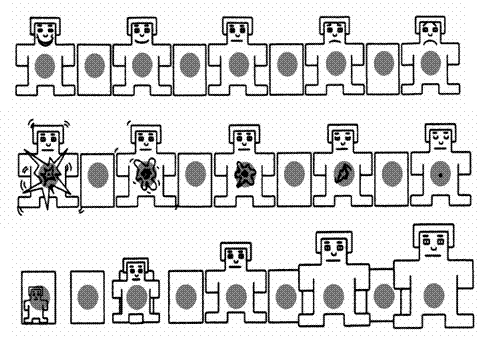
Figura 1. Escalas del Self-Assessment Manikin (SAM): Valencia (fila
superior), Arousal (fila intermedia) y Dominancia (fila inferior).
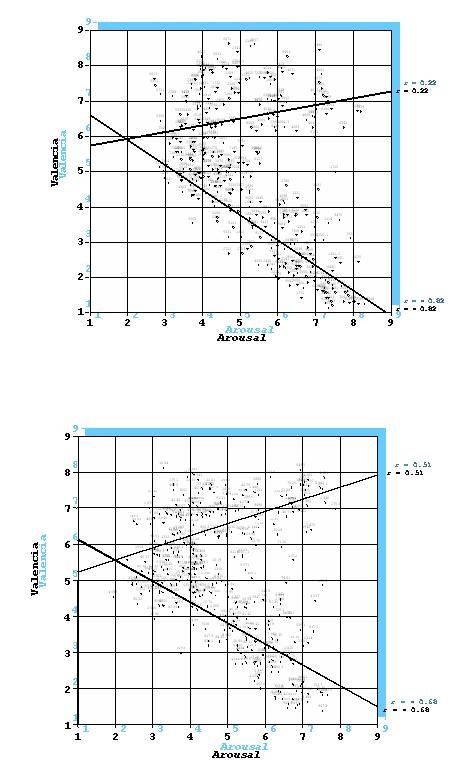
Figura 2. Representación de las imágenes del IAPS en el espacio bidimendional Valencia-Arousal para mujeres (gráfica superior) y varones (gráfica inferior)
Las evaluaciones afectivas de las imágenes del IAPS han demostrado ser altamente fiables y estables. La figura 2 representa la distribución de las últimas 352 fotografías del IAPS en el espacio bidimensional definido por la valencia y el arousal, arriba para el grupo de varones y abajo para el grupo de mujeres. El eje vertical codifica la posición de cada fotografía en la dimensión de valencia (1= altamente desagradable, 5 = neutra, 9 = altamente agradable). El eje horizontal codifica su posición en la dimensión de arousal (1 = nada activadora, 5 = moderadamente activadora, 9 = altamente activadora). Cada punto entre los dos ejes representa la media de las estimaciones de cada una de las fotografías. Como puede verse, la distribución de las imágenes adopta la forma de boomerang, con dos brazos que salen desde una zona afectivamente neutra y mínima activación hacia los dos extremos agradable y desagradable de máxima activación. Estos resultados reproducen los obtenidos con las primeras 480 fotografías del IAPS y con los obtenidos en las muestras estadounidenses. Además, se aprecia que el brazo que se extiende hacia el polo agradable tiene una menor inclinación, junto con una mayor dispersión, que el brazo que se extiende hacia el polo desagradable. También se observa la práctica ausencia de imágenes en algunas regiones del espacio bidimensional, en particular en el cuadrante poco activador y desagradable y en el cuadrante altamente activador y neutro.
Estos datos tienen claras implicaciones teóricas. Por una parte, desconfirman el llamado modelo circular o circumplejo propuesto por algunos autores (Russell, 1980), al menos en lo que respecta a estímulos afectivos perceptuales. Por otra parte, la diferente inclinación de la distribución de las imágenes en función de la valencia hedónica –mayor inclinación para las imágenes desagradables que para las imágenes agradables- sugiere mecanismos de acción diferente para los sistemas motivacionales apetitivo y defensivo. La menor inclinación para el sistema apetitivo indicaría la presencia de un sesgo hacia la positividad cuando ninguno de los dos sistemas motivacionales está fuertemente activado. En otras palabras, cuando los niveles motivacionales son bajos (poca activación), la tendencia a la aproximación sería mayor que la tendencia a la evitación, lo que explicaría las reacciones exploratorias y de orientación típicas de la vida cotidiana, cuando los estímulos ambientales no son ni fuertemente amenazantes ni fuertemente atractivos. Por el contrario, la mayor inclinación para el sistema defensivo indicaría un sesgo hacia la negatividad cuando los niveles motivacionales son muy altos, esto es, habría una mayor tendencia a responder con reacciones defensivas de evitación y escape que con reacciones positivas de aproximación cuando aumentan los niveles de activación emocional. Este sesgo de negatividad o conservadurismo adaptativo, como lo ha denominado Mineka (1992), tiene claras funciones de cara a la supervivencia, debido al coste potencialmente mortal de la ausencia de dicho sesgo.
Pese a la similitud de los baremos españoles y estadounidenses, así como de los resultados generales de las muestras de varones y mujeres, existen también importantes diferencias que apelan a la necesidad de utilizar datos normativos adecuados al tipo de persona y al contexto cultural de cada investigación. Las diferencias entre personas españolas y estadounidenses no se producen en las evaluaciones de la dimensión de valencia, pero sí en las dimensiones del arousal y la dominancia: las españolas perciben las imágenes con un mayor nivel de activación y con un menor nivel de dominio o control que los estadounidenses. Estos resultados sugieren que las personas españolas reaccionan emocionalmente con mayor intensidad, pero con menor control, que las estadounidenses, algo que encaja bastante bien con los estereotipos que existen sobre ambas poblaciones. Así mismo, existen diferencias de género entre varones y mujeres. Como puede apreciarse en la figura 3, el brazo del boomerang que se extiende hacia el polo desagradable tiene una mayor inclinación, junto con una menor dispersión, en el caso de las mujeres, mientras que el brazo que se extiende hacia el polo agradable tiene una mayor inclinación, junto con una menor dispersión, en el caso de los varones. Las diferencias de género son también evidentes cuando se identifican las imágenes evaluadas por varones y mujeres como más agradables. Las mujeres asignan puntuaciones mayores a las imágenes de bebés, familias, atracciones de feria y parejas románticas, mientras que los varones consideran como las más agradables las imágenes eróticas y de deportes.
En general, las imágenes del IAPS proporcionan apoyo empírico a la idea de que las emociones se organizan jerárquicamente en torno a dos sistemas motivacionales primarios, el apetitivo y el defensivo, además de proporcionar un método de inducción de estados emocionales mediante el uso de estímulos calibrados cuantitativamente y adaptados al contexto sociocultural en el que se van a aplicar. Su uso en las investigaciones de laboratorio está permitiendo avanzar en la tarea de desentrañar las complejas relaciones existentes entre los tres sistemas de respuesta emocional –el conductual, el expresivo-lingüístico y el fisiológico-, así como en el conocimiento de los mecanismos psicológicos y fisiológicos que controlan nuestras emociones.
4.3 La modulación emocional del reflejo de sobresalto
El paradigma de visualización de imágenes afectivas ha desvelado la existencia de covariaciones sistemáticas entre determinados componentes de las reacciones emocionales. La dimensión subjetiva de valencia muestra una clara covariación con diversas respuestas fisiológicas: el reflejo de sobresalto, la frecuencia cardíaca y la actividad electromiográfica de los músculos corrugador y cigomático (responsables del fruncimiento de las cejas y el estiramiento de los labios, respectivamente). Las imágenes desagradables producen mayor amplitud del reflejo de sobresalto, mayor desaceleración cardíaca, mayor tensión en el músculo corrugador y menor tensión en el músculo cigomático, que las imágenes neutras y agradables, en este orden. La dimensión subjetiva de arousal, por su parte, muestra una clara covariación con la conductancia eléctrica de la piel, los potenciales corticales evocados, el tiempo de visión de las imágenes, la evaluación subjetiva de interés por las imágenes y las medidas de recuerdo y reconocimiento de imágenes previamente presentadas. Las imágenes más activantes –tanto agradables como desagradables- producen cambios mayores en todas estas medidas que las imágenes neutras –las menos activantes. Cuando se aplica la metodología factorial a este conjunto de datos expresivo-lingüísticos, fisiológicos y conductuales aparece una estructura factorial con dos factores: un primer factor, que explica la mayor parte de la varianza, con altas saturaciones en valencia subjetiva, reflejo de sobresalto, tasa cardíaca y actividad electromiográfica del corrugador y cigomático, y un segundo factor con altas saturaciones en arousal subjetivo, evaluaciones de interés por las imágenes, tiempo de visión, conductancia de la piel y potenciales corticales. Estos datos vuelven a reforzar la idea de que los afectos se construyen jerárquicamente en torno a determinantes motivacionales relacionados con los sistemas apetitivo y defensivo (Bradley, 2000).
De todas las medidas fisiológicas y conductuales que covarían con las dimensiones emocionales, la del reflejo de sobresalto ha sido, sin duda, la que mayor interés ha despertado entre los investigadores. La modulación de la amplitud del reflejo de sobresalto cuando las personas visualizan imágenes de diferente contenido afectivo es uno de los fenómenos más robustos encontrados en la investigación experimental. El reflejo de sobresalto es un patrón de activación motora elicitado por estimulación intensa o aversiva de inicio repentino. Fue descrito por Landis y Hunt (1939) como una acción refleja con tres componentes: (a) un parpadeo, (b) una inclinación brusca de la cabeza hacia delante, y (c) una onda de flexión descendente que se transmite desde el tronco hasta las rodillas. Existe una amplia literatura sobre la modulación emocional de este reflejo tanto en animales (Davis, 1992; LeDoux, 1996, 2003) como en humanos (Lang, 1995). La medida más utilizada del reflejo de sobresalto en humanos se basa en el registro psicofisiológico del parpadeo –el componente primero y más estable del sobresalto- a través de la electromiografía (EMG) del músculo orbicular del ojo.
El reflejo puede ser elicitado por diferentes estímulos: soplo de aire, luces intensas, ruidos repentinos o pequeños calambres. La modulación del reflejo de sobresalto por factores emocionales fue descrita por primera vez por Brown, Kalish y Farber (1951) en el contexto de las investigaciones del grupo de Yale sobre el miedo condicionado en ratas: la respuesta de sobresalto, medida mediante un estabilímetro localizado en el suelo de la caja de experimentación, fue mayor cuando el estímulo provocador del sobresalto –el disparo de una pistola de juguete- se presentaba simultáneamente con señales previamente condicionadas de miedo. Los primeros estudios sobre la modulación del reflejo de sobresalto en humanos fueron de los de Vrana, Spence y Lang (1988). Estos autores utilizaron el paradigma de la visualización de imágenes afectivas, superpuesto a la prueba de sobresalto, en sustitución de los estímulos condicionados de miedo utilizados en los estudios con animales. Este paradigma implica la presentación del estímulo provocador del sobresalto –generalmente un ruido blanco de 50 milisegundos de duración, 95 dB de intensidad y tiempo de subida o risetime instantáneo, presentado a través de auriculares- en un contexto de percepción visual de imágenes afectivas seleccionadas del IAPS y proyectadas delante de la persona durante unos segundos. En estos estudios se encontró que la magnitud del reflejo variaba sistemáticamente en función del contenido afectivo de las diapositivas: la magnitud del reflejo de sobresalto era máxima durante la exposición de diapositivas desagradables y mínima durante la exposición de diapositivas agradables, quedando en una magnitud intermedia durante la exposición de diapositivas neutras.
Lang ha explicado este efecto proponiendo la hipótesis del priming (facilitación) motivacional. La hipótesis predice que se observará potenciación de un reflejo cuando exista congruencia entre el tipo de reflejo (defensivo o apetitivo) y el estado motivacional propio del estado afectivo en que se encuentre el organismo (defensivo o apetitivo). Por el contrario, se observará inhibición del reflejo cuando exista incongruencia entre el tipo de reflejo y el estado motivacional en que se encuentre el organismo. Desde esta perspectiva, se considera que el reflejo de sobresalto es un reflejo intrínsecamente protector o defensivo propio del sistema motivacional aversivo. Los estímulos desagradables (por ejemplo, las diapositivas de cuerpos mutilados) inducen un estado emocional negativo, también propio del sistema motivacional aversivo. La congruencia entre ambos estados provoca la potenciación del reflejo. Por el contrario, los estímulos agradables (por ejemplo, diapositivas de contenido sexual), al inducir un estado emocional positivo, propio del sistema motivacional apetitivo y, por tanto, opuesto al sistema motivacional activado por el estímulo del sobresalto, produciría la inhibición del reflejo.
5. Regulación emocional: entre el corazón y el cerebro
5.1 La respuesta cardíaca de defensa
El modelo del priming motivacional es aplicable a todos los reflejos defensivos y no sólo al reflejo motor de sobresalto. La respuesta cardíaca de defensa es otro tipo de respuesta defensiva del organismo con una larga tradición en la investigación psicofisiológica desde la época de James y Cannon. Cannon la identificó con la respuesta de lucha o huida y, por tanto, con una clara significación motivacional de preparación para acciones adaptativas ante situaciones de amenaza o peligro. La investigación reciente sobre la respuesta cardíaca de defensa se ha llevado a cabo utilizando procedimientos similares a los utilizados en el estudio del reflejo motor de sobresalto: presentación de estímulos auditivos de alta intensidad. El estímulo elicitador ha sido generalmente un ruido blanco de 100-110 dB de intensidad, entre 500 y 1000 milisegundos de duración y tiempo de subida instantáneo. El registro de la respuesta se realiza a partir del registro del electrocardiograma analizando los cambios en la tasa cardíaca latido a latido durante el período inmediatamente posterior a la presentación del estímulo.
La figura 3 recoge un típico registro de la respuesta cardíaca de defensa ante tres presentaciones de un estímulo auditivo (la tres líneas verticales discontinuas) con una duración total del registro de 5 minutos. El canal superior corresponde a la respiración, el canal intermedio a la amplitud del pulso digital y el inferior a la tasa cardíaca. Como puede verse en el canal inferior, la respuesta cardíaca ante la primera presentación del estímulo no es una simple aceleración cardíaca. Es un patrón complejo de cambios acelerativos y desacelerativos secuenciales que tienen lugar dentro de los 80-90 segundos posteriores al estímulo: una primera aceleración con amplitud máxima en torno al segundo 3, una desaceleración inmediatemente posterior, una segunda aceleración mucho más prolongada que alcanza su amplitud máxima en torno al segundo 35, y una lenta desaceleración final que se estabiliza en torno a los 80 segundos. El patrón completo de la respuesta se observa prácticamente sólo en la primera presentación del estímulo, produciéndose una rápida habituación de la respuesta que afecta sobre todo al segundo componente acelerativo.
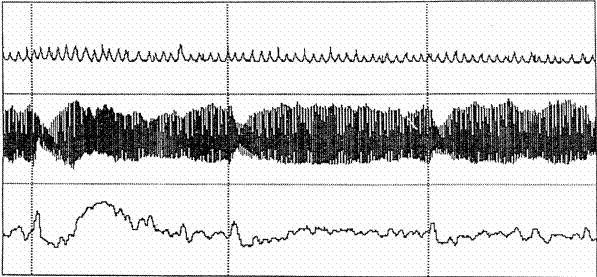
Figura 3. Registro de la Respuesta Cardíaca de Defensa (línea inferior) ante tres presentaciones de un ruido intenso (líneas verticales).
Los impresionantes cambios rítmicos que se producen en el corazón de forma secuencial ante un ruido intenso inesperado son la consecuencia de una compleja intervención de mecanismos neurofisiológicos simpáticos y parasimpáticos activados desde diversos núcleos del tronco cerebral y sobre los que actúan estructuras cerebrales superiores implicadas en la regulación emocional. Diferentes estudios (Reyes del Paso, Godoy y Vila, 1993; Reyes del Paso, Vila y García, 1994) han demostrado que la primera aceleración y primera desaceleración están controladas vagalmente: la primera aceleración respondería a una inmediata inhibición vagal, mientras que la desaceleración posterior respondería a un incremento subsecuente de la activación vagal. En cuanto a la segunda aceleración y segunda desaceleracion, los mecanismos implicados son fundamentalmente simpáticos y parasimpáticos actuando recíprocamente: aumento de la activación simpática acompañada de disminución de la activación parasimpática durante la segunda aceleración, y disminución de la activación simpática acompañada de activación parasimpática durante la segunda desaceleración (Fernández y Vila, 1989). Estos mecanismos están regulados por una red compleja de estructuras cerebrales que van desde áreas prefrontales hasta el tronco del cerebro pasando por el núcleo central de la amígdala (Thayer y Siegle, 2002). De acuerdo con la hipótesis del priming motivacional, los mismos mecanismos que regulan la modulación emocional del reflejo motor de sobresalto deberían participar en la regulación de la respuesta cardíaca de defensa. De ahí la importancia de investigar simultáneamente ambos reflejos. La convergencia de datos procedentes del estudio de dos respuestas defensivas diferentes consolidaría los modelos teóricos propuestos y permitiría avanzar en el conocimiento de los mecanismos neurofisiológicos de la emoción.
El estudio simultaneo del reflejo de sobresalto y de la respuesta cardíaca de defensa requiere el uso de una metodología común. Varios estudios han demostrado que manipulando simultáneamente la intensidad, la duración y el tiempo de subida o risetime del estímulo elicitador podemos provocar simultáneamente el reflejo motor de sobresalto y la respuesta cardíaca de defensa, sólo el reflejo motor de sobresalto o sólo la respuesta cardíaca de defensa. También se ha podido descartar que en el patrón de la respuesta cardíaca de defensa existan dos componentes diferenciados, uno de sobresalto (relacionado con la primera aceleración) y un componente de defensa propiamente dicho (relacionado con la segunda aceleración). Ambos componentes acelerativos, junto con sus dos componentes desacelerativos, forman parte del patrón secuencial de la respuesta cardíaca de defensa, dado que cuando se utilizan tiempos de subida que descartan el sobresalto, continúa produciéndose el patrón completo de la respuesta.
5.2 Modulación emocional de reflejos defensivos: sobresalto y defensa
La investigación simultanea de la modulación emocional del sobresalto motor y la defensa cardíaca, utilizando el procedimiento de visualización de imágenes, se ha llevado a cabo en varios estudios. Resumimos aquí dos estudios en los que la visualización de las imágenes se realizó bajo condiciones de procesamiento consciente. En el primero de ellos (Sánchez, Ruiz-Padial, Pérez, Fernández, Cobos y Vila, 2002), un grupo de estudiantes voluntarias pasaron la prueba de visualización de 9 diapositivas afectivas, en orden balanceado, seleccionadas del IAPS español: 3 agradables, 3 neutras y 3 desagradables. El estímulo evocador de los dos reflejos defensivos (ruido blanco intenso, tiempo de subida instantáneo y medio segundo de duración) se presentaba a través de auriculares a los tres segundos y medio de iniciarse la proyección de las 9 diapositivas.
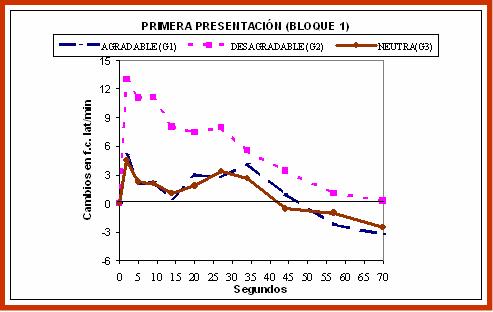
Figura 4. Respuesta Cardíaca de Defensa durante la visualización de imágenes afectivas (agradable, neutra y desagradable).
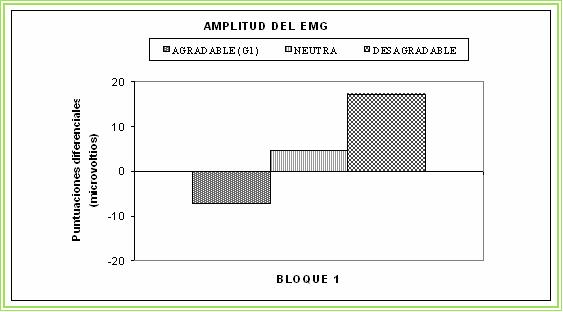
Figura 5. Reflejo Motor de
Sobresalto durante la visualición de imágenes afectivas (agradables, neutras
y desagradables).
En la figura 4, presentamos los resultados referidos a la respuesta cardíaca de defensa ante el estímulo evocador de los reflejos defensivos durante la visualización de la primera diapositiva de cada categoría emocional. Como puede verse, la respuesta cardíaca cuando se visualiza la diapositiva desagradable queda muy afectada, desapareciendo el patrón típico aceleración-desaceleración-aceleración-desaceleración. En su lugar, se encuentra una única aceleración muy aumentada en la que parecen unirse los dos componentes acelerativos de la respuesta, haciendo desaparecer totalmente la primera desaceleración. En el caso de la visualización de las diapositivas agradable y neutra, la respuesta cardíaca se parece más al patrón típico excepto que todos los componentes están ligeramente disminuidos. Los resultados relativos al reflejo motor de sobresalto se presentan en la figura 5. Como puede verse, se reproduce la esperada modulación del reflejo motor de sobresalto: la amplitud del reflejo se potencia cuando se está visualizando la diapositiva desagradable (valor por encima de la media) y se inhibe cuando se está visualizando la diapositiva agradable (valor por debajo de la media), quedando la diapositiva neutra en una posición intermedia.
El segundo estudio (Vila y col., 2003) se realizó seleccionando otro conjunto de estudiantes voluntarias en función de sus puntuaciones en tres cuestionarios específicos de fobias: uno de fobia a las serpientes, otro de fobia a las arañas y el otro de fobia a la sangre-mutilaciones. Las participantes se distribuyeron en tres subgrupos: subgrupo de miedo a los animales (serpiente o araña) pero sin miedo a la sangre-mutilaciones; subgrupo de miedo a la sangre-mutilaciones pero sin miedo a las serpientes o arañas; y subgrupo sin miedo a los animales ni a la sangre-mutilizaciones. Todas las participantes pasaron por una prueba de visualización de dos imágenes seleccionadas del IAPS español: una de sangre y la otra del animal fóbico (serpiente o araña). Siguiendo un procedimiento similar al del estudio anterior, las dos diapositivas se proyectaron durante 6 segundos, balanceando el orden de presentación. El estímulo evocador de ambos reflejos se presentaba a los tres segundos y medio de iniciarse la diapositiva. Los resultados volvieron a mostrar una respuesta cardíaca muy potenciada cuando las participantes visualizaban la imagen fóbica frente a la imagen no fóbica. La forma de la respuesta ante la imagen fóbica volvía a reproducir los resultados del estudio anterior: desaparición del primer componente desacelerativo y fusión incrementada de los dos componentes acelerativos. En cuanto al reflejo motor de sobresalto, los resultados vuelven a confirmar la hipótesis del priming motivacional: el grupo con fobia a los animales mostró una mayor amplitud del sobresalto ante la diapositiva del animal que ante la diapositiva de sangre; el grupo con fobia a la sangre mostró una mayor amplitud ante la diapositiva de sangre que ante la diapositiva del animal; mientras que el grupo no fóbico mostró una respuesta disminuida ante ambas diapositivas.
Los resultados de estos dos estudios coinciden en poner de manifiesto un importante efecto modulador del contenido afectivo de las imágenes tanto sobre la respuesta cardíaca de defensa como sobre el reflejo motor de sobresalto. En el caso de la respuesta cardíaca de defensa, nuestros resultados muestran no sólo una marcada potenciación de los componentes acelerativos de la respuesta ante las imágenes desagradables y fóbicas, reforzando el modelo del priming motivacional, sino también una marcada modificación de la topografía de la respuesta: la primera y segunda aceleración parecen juntarse adelantándose la segunda aceleración y desapareciendo completamente la primera desaceleración. Esta modificación del patrón de la respuesta en función de la presentación previa de estímulos afectivos sugiere la presencia de mecanismos psicológicos y fisiológicos mediadores de las reacciones defensivas que pueden ser clave para entender el funcionamiento del cerebro y su papel en la regulación emocional.
Cuando el estímulo auditivo intenso se presenta de forma inesperada, esto es, sin señales de aviso o estímulos de priming, el patrón de cambios acelerativos y desacelerativos refleja una secuencia de procesos psicológicos de tipo atencional y emocional, coherente con las reacciones defensivas descritas en animales cuando se encuentran en contextos naturales (por ejemplo, ante la presencia de un depredador). En tales contextos, las primeras fases de la reacción defensiva –coincidentes con la primera aceleración y primera desaceleración mediadas vagalmente- están caracterizadas por un predominio de los procesos atencionales dirigidos al análisis del estímulo aversivo, mientras que las fases siguientes –coincidentes con la segunda aceleración y segunda desaceleración mediadas simpáticamente- están caracterizadas por un predominio de los procesos motivacionales dirigidos a la preparación de acciones defensivas de lucha o huida. Sin embargo, esta secuencia parece alterarse cuando el estímulo aversivo está precedido de estímulos de priming -la visualización de imágenes desagradables y fóbicas u otras señales de aviso. En este caso, la secuencia parece adelantarse temporalmente: las primeras fases atencionales pasan a los estímulos previos y las fases finales preparatorias de las acciones defensivas ocupan el núcleo central de la respuesta cardíaca al estímulo aversivo.
5.3 Modulación preatencional de reflejos defensivos: la no consciencia
La investigación del procesamiento no consciente de estímulos tiene una gran relevancia para la psicología y, sobre todo, para la psicología clínica. El procesamiento no consciente de estímulos emocionales permite entender muchos de los comportamientos patológicos encontrados en personas con trastornos de ansiedad o con trastornos neuropsicológicos. Además, el tema de la no consciencia conecta con el núcleo de las explicaciones psicoanalíticas de la patología: el inconsciente. Una de las líneas de investigación más sólidas sobre el procesamiento no consciente de estímulos emocionales es la desarrollada por el grupo de Arne Öhman en la universidad de Uppsala y en el Instituto Karolinska de Suecia. El eje central de las investigaciones de Öhman es la consideración de las emociones desde un punto de vista evolucionista. Öhman analiza el miedo dentro de dos sistemas conductuales desarrollados a lo largo de la evolución: un sistema de defensa del depredador y un sistema de sumisión social. El sistema de defensa del depredador explicaría el miedo a los animales, se habría desarrollado a partir de un miedo a los reptiles en los primeros mamíferos, y promovería un rápido escape de depredadores potenciales. Por su parte, el sistema de dominancia-sumisión daría lugar a los miedos sociales, siendo su función adaptativa el promover el orden social facilitando el establecimiento de jerarquías de dominancia. El argumento básico es que la evolución ha dotado a los humanos con una predisposición a asociar miedo con situaciones que amenazaron la supervivencia de nuestros antepasados.
Los datos que apoyan esta idea proceden de tres tipos de estudios experimentales (Öhman y Mineka, 2001): en primer lugar, estudios sobre condicionamiento clásico preparado del miedo en humanos utilizando estímulos biológicamente relevantes (serpientes, arañas, rostros amenazantes) frente a estímulos biológicamente irrelevantes (flores, paisajes, rostros alegres); en segundo lugar, estudios sobre aprendizaje vicario u observacional del miedo en monos rhesus, utilizando videos de otros monos reaccionando con miedo o sin miedo ante estímulos con los que no habían tenido experiencia previa los observadores: cocodrilos y serpientes (biológicamente relevantes) frente a flores y conejos (biológicamente irrelevantes); en tercer lugar, estudios sobre procesamiento no consciente de estímulos amenazantes utilizando la técnica de enmascaramiento hacia atrás. Esta técnica consiste en impedir el reconocimiento del estímulo amenazante mediante su presentación seguida de otro estímulo-máscara (formado por trocitos recortados y unidos al azar de las imágenes amenazantes) con un intervalo inferior a 50 milisegundos (Esteves, Dimberg y Öhman, 1994). Bajo estas condiciones, el estímulo-máscara bloquea el reconocimiento del estímulo amenazante. No obstante, se puede demostrar que es procesado por el sistema, sin requerir atención consciente, y que influye en la conducta emocional de la persona.
En una serie elegante de estudios, el grupo de Öhman ha demostrado (a) que se pueden elicitar respuestas psicofisiológicas de miedo ante estímulos amenazantes (rostros enfadados frente a rostros felices, y serpientes y arañas frente a flores y setas) no percibidos conscientemente, (b) que se pueden elicitar respuestas psicofisiológicas de miedo, previamente condicionadas a estímulos biológicamente preparados bajo condiciones conscientes, aunque estos estímulos se presenten posteriormente de forma enmascarada, (c) que se pueden condicionar respuestas psicofisiológicas de miedo a estímulos biológicamente preparados presentados durante el condicionamiento de forma enmascarada, y (d) que existen correlaciones entre las respuestas psicofisiológicas y las evaluaciones subjetivas: ante las imágenes amenazantes enmascaradas las personas se evalúan a sí mismos como más a disgusto, más activados y con menor control que ante las imágenes no amenazantes enmascaradas. Estos datos permiten explicar que, en determinados trastornos de ansiedad, el miedo se experimente como irracional e incontrolable. La activación autonómica, que constituye un componente central en tales trastornos, puede estar actuando antes de que la persona sea consciente de los estímulos amenazantes. Dicha activación fisiológica podría sesgar el análisis posterior de los estímulos, cuando estos son procesados conscientemente, en el sentido de sobrevalorar la amenaza y el miedo que provocan. Desde este punto de vista, no es sorprendente, por ejemplo, que las personas con miedos fóbicos se sientan incapaces de controlar su miedo y centren sus estrategias de afrontamiento en el escape y la evitación.
Los estudios del grupo de Öhman han utilizado el reflejo de orientación, medido a través de la conductancia eléctrica de la piel, como la principal variable indicadora del procesamiento no consciente de los estímulos amenazantes. Esta medida psicofisiológica es altamente sensible a procesos atencionales y emocionales. Sin embargo, tiene el inconveniente de no permitir la diferenciación entre orientación y defensa. Aunque Öhman interpreta sus datos en términos de reacciones emocionales de miedo y, por tanto, como reacciones defensivas, cabe la posibilidad de una interpretación alternativa en términos puramente atencionales o pre-atencionales de orientación a los estímulos. Obtener evidencia inequívoca de la activación del sistema defensivo durante el procesamiento no consciente de los estímulos amenazantes daría un apoyo adicional al modelo teórico de Öhman. En este sentido, tanto el reflejo motor de sobresalto como la respuesta cardíaca de defensa podrían aportar datos clarificadores de los mecanismos activados durante el procesamiento no consciente de la amenaza. Para ello adaptamos el procedimiento de visualización de imágenes afectivas, descrito en el apartado anterior, a la técnica de enmascaramiento hacia atrás utilizada por Öhman y realizamos una serie de estudios para examinar si la modulación emocional de los reflejos defensivos, demostrada bajo condiciones conscientes, se produce igualmente bajo condiciones no conscientes.
En un primer estudio (Ruiz-Padial, Sánchez, Thayer y Vila, 2002), seleccionamos un grupo de estudiantes voluntarias con miedo intenso a las arañas y las dividimos en dos subgrupos: un grupo que pasó la tarea de visualización de las imágenes bajo condiciones de enmascaramiento no efectivo (grupo consciente) y un grupo que pasó la tarea bajo condiciones de enmascaramiento efectivo (grupo no consciente). Cada ensayo consistía en una secuencia de seis presentaciones de la imagen (una araña o una flor) seguida de la máscara a un ritmo de una presentación por segundo. El estímulo evocador de la respuesta cardíaca de defensa y el reflejo motor de sobresalto –un ruido blanco similar al de los estudios anteriores- aparecía hacia la mitad de la secuencia coincidiendo con la cuarta presentación de la máscara. Una vez finalizada la tarea, las participantes realizaban una prueba de reconocimiento de los estímulos. Se les presentaba una lámina con 12 imágenes (4 de arañas, 4 de flores y 4 de serpientes, incluidas las 2 utilizadas durante la tarea) y tenían que indicar las dos imágenes que se habían presentado bien bajo condiciones conscientes bien bajo condiciones no conscientes.
La figura 6 muestra los resultados obtenidos. Tanto el grupo consciente como el no consciente mostraron la potenciación de la respuesta cardíaca de defensa cuando visualizaban la imagen fóbica en comparación con la imagen no fóbica, siendo el efecto más pronunciado en el grupo consciente. El reflejo de sobresalto también mostró el mismo efecto, observándose al mismo tiempo un efecto de sensibilización: la amplitud del sobresalto fue mayor en la segunda presentación del estímulo auditivo. En este ensayo es en el que se observa claramente la potenciación del reflejo ante la visualización de la imagen fóbica en ambos grupos. En cuanto a la forma de la respuesta cardíaca de defensa, se vuelve a reproducir la ausencia de la primera desaceleración lo que sugiere que la secuencia defensiva, temporalmente adelantada cuando el estímulo defensivo va precedido de imágenes desagradables o fóbicas, se produce también bajo condiciones de procesamiento no consciente.
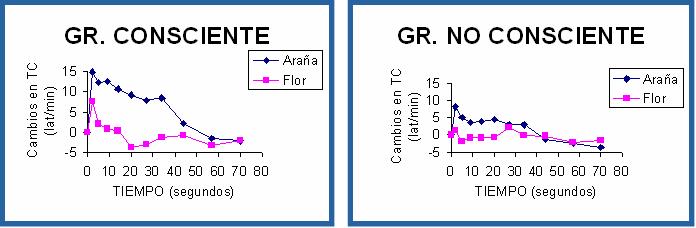
Figura 6. Respuesta Cardíaca de Defensa (arriba) y Reflejo Motor de Sobresalto (abajo) durante la visualización de la imagen fóbica y no fóbica en los grupos consciente (izquierda) y no consciente (derecha).
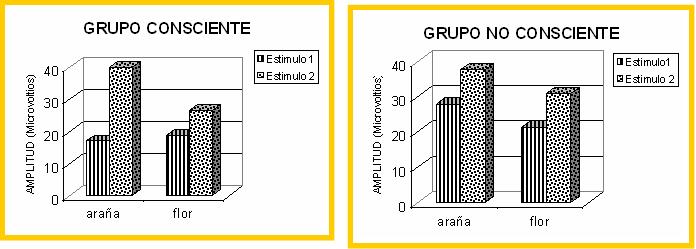
En un segundo estudio (Ruiz-Padial, 2003), se seleccionó otro grupo de estudiantes voluntarias, la mitad con miedo intenso a las serpientes o las arañas pero sin miedo a la sangre-mutilaciones y la otra mitad con miedo intenso a la sangre-mutilaciones pero sin miedo a las serpientes o las arañas. Todas las participantes pasaron por una tarea sólo de sobresalto (ruido blanco intenso de 50 milisegundos de duración y tiempo de subida instantáneo) durante la visualización, en orden aleatorio, de 20 imágenes seleccionadas del IAPS español: 5 agradables, 5 neutras, 5 de sangre y 5 de animal (serpiente o araña). La mitad de las participantes en cada grupo pasaron la tarea bajo condiciones de enmascaramiento efectivo (grupo no consciente) y la otra mitad bajo condiciones de enmascaramiento no efectivo (grupo consciente). Cada ensayo de visualización consistía en una única presentación de la imagen seguida de la máscara. Finalizada la tarea, las participantes pasaban una prueba de reconocimiento de las imágenes en la que se presentaban, en varias láminas, 40 imágenes -las 20 visualizadas durante la tarea y 20 imágenes distractoras (5 de cada contenido)- teniendo que indicar las participantes qué imágenes se habían presentado y con qué grado de seguridad emitían su juicio.
Los resultados de este estudio volvieron a confirmar el efecto modulador de las imágenes fóbicas tanto en el grupo consciente como en el grupo no consciente y tanto en la respuesta cardíaca de defensa como en el reflejo motor de sobresalto. Los resultados de estos dos estudios, por tanto, ofrecen apoyo empírico a la idea de la existencia de procesamiento no consciente de estímulos visuales capaces de activar el sistema motivacional defensivo. La capacidad de estos estímulos de ser procesados no conscientemente y de potenciar o inhibir las reacciones defensivas implica que existen vías directas de activación del circuito neural defensivo -fundamentalmente la amígdala- que no pasan por las estructuras corticales propias del procesamiento consciente (LeDoux, 1996). La investigación neurofisiológica en humanos utilizando técnicas de neuroimagen –resonancia magnética funcional y tomografía por emisión de positrones- también confirma la vía directa de activación de la amígdala durante el procesamiento no consciente de estímulos emocionales. Se ha demostrado, por ejemplo, que la presentación enmascarada de rostros enfadados produce incrementos en la actividad de la amígdala mientras que la presentación también enmascarada de rostros felices produce disminuciones (Morris, Öhman y Dolan, 1998, 1999; Whalen y colaboradores, 1998). Los centros subcorticales que regulan las emociones pueden, por tanto, activarse o inhibirse en presencia de señales de peligro o de seguridad sin requerir procesamiento consciente.
6. Implicaciones teóricas
La investigación experimental sobre la emoción desde la nueva perspectiva psicofisiológica tiene importantes implicaciones teóricas tanto con respecto a las emociones y motivaciones relacionadas con el sistema defensivo (miedo, ansiedad, ira, tristeza, estrés, dolor, sufrimiento ) como con respecto al estudio científico de la emoción, en general.
Con respecto al sistema defensivo, la principal implicación es la reorientación del estudio de los procesos protectores del organismo hacia el contexto natural de las reacciones defensivas. Cuando las reacciones defensivas se estudian en contextos relevantes para la supervivencia de los organismos, como ante la inminencia de un ataque por un potencial depredador o un delincuente, es evidente que la reacción defensiva no es una respuesta única que se pueda conceptuar como cognitiva frente a emocional, o viceversa. La reacción defensiva sigue un proceso secuencial dinámico con fases iniciales en las que predominan factores atencionales motivados aversivamente, dirigidos a la detección y análisis del peligro potencial, y fases posteriores en las que predominan las acciones defensivas de lucha o huida. Dependiendo de la proximidad espacial y temporal del peligro, diferentes componentes de la reacción defensiva irían apareciendo de forma sucesiva. En este sentido, el patrón de la respuesta cardíaca de defensa, con componentes acelerativos y desacelerativos, y con significación tanto atencional como emocional, debe entenderse como el reflejo de la estrecha relación existente entre procesos cognitivos favorecedores del análisis del peligro y procesos motivacionales favorecedores de la subsecuente acción adaptativa. Se trata de un proceso dinámico en el que los componentes pueden adelantarse, retrasarse, potenciarse o inhibirse, en función del estado emocional previo del organismo y del éxito o fracaso de las transiciones entre sus diferentes componentes.
Desde este planteamiento procesual, puede también entenderse el carácter dinámico del concepto de estrés y su estrecha relación con el concepto de defensa. El estrés no sería una respuesta defensiva concreta, sería el estado mantenido de activación del sistema motivacional defensivo, un estado que favorece la continua aparición de respuestas defensivas diversas, en función del contexto, y cuyos indicadores psicofisiológicos correlacionan con una pobre salud física y psicológica: incrementos en la tasa cardíaca mediada simpáticamente y decrementos en la variabilidad cardíaca mediada vagalmente (arritmia sinusal respiratoria y reflejo barorreceptor).
La investigación sobre la modulación de los reflejos defensivos también tiene importantes implicaciones con respecto al estudio científico de la emoción. Las viejas dicotomías periferalismo-centralismo, especificidad-dimensionalidad y fisiología-cognición parecen perder sentido. Con respecto al debate periferalismo-centralismo, los datos claramente apuntan a que el componente subjetivo de las emociones no se deteriora cuando personas adultas pierden parte de su feedback periférico como consecuencia de lesiones o enfermedades graves (Birbaumer, 2001; Cobos y col., 2003). Pero estos mismos datos dejan también claro que en estas personas las respuestas fisiológicas periféricas siguen mostrando el mismo tipo de modulación emocional que se observa en personas sin tales restricciones periféricas. Las interacciones centro-periferia están inmersas en el propio funcionamiento del cerebro, como ha señalado LeDoux (1996, 2003).
Lo mismo podemos decir con respecto al debate especificidad-dimensionalidad y fisiología-cognición. La vieja idea de que la fisiología periférica es inespecífica con respecto a las emociones contribuyendo sólo a la intensidad emocional fue desmontada en los años 60 con la misma contundencia con la que Cannon intentó desmontar cuarenta años antes la idea contraria defendida por James (Lacey, 1967). La especificidad se encuentra incluso en las grandes dimensiones emocionales –valencia y arousal- tal como hemos expuesto al hablar de las correlaciones entre los indicadores fisiológicos y subjetivos del procesamiento de imágenes afectivas. Especificidad y dimensionalidad están, por tanto, igualmente inmersas en la organización estructural y funcional del cerebro. Por último, el debate fisiología-cognición -o emoción versus cognición- también deja de tener sentido. Del mismo modo que es un error reducir la emoción a un proceso cognitivo, es un error negar la implicación de determinados procesos cognitivos –en especial, la atención y la memoria- en la regulación emocional. Las conexiones de las áreas subcorticales propias del procesamiento emocional –entre ellas, la amígdala- con las áreas corticales y subcorticales propias del procesamiento cognitivo –entre ellas, la corteza prefrontal y el hipocampo- son tan numerosas que es imposible que no se produzcan interacciones entre ambas. Es lo que explica que muchas reacciones emocionales se puedan disparar por la simple anticipación de un peligro o por el recuerdo explícito de un suceso emocional. O que una vez disparada una reacción emocional por la vía directa –no consciente- ésta pueda ser modulada y controlada por determinadas acciones o pensamientos de los que sí somos conscientes y que podemos aprender a utilizar con eficacia.
El estudio de la emoción desde la perspectiva psicofisiológica que hemos expuesto en este trabajo defiende un marco teórico integrador: la emoción implica procesos que son simultáneamente periféricos y centrales, específicos y dimensionales, y fisiológicos y cognitivos. Desde este marco se sugiere la conveniencia de abandonar definitivamente las tradicionales dicotomías y controversias que han caracterizado el estudio científico de la emoción desde la época de James y Cannon.
REFERENCIAS
Bower, G. H. (1981). Mood and memory. American Psychologist, 36, 129-148.
Bradley, M. M. (2000). Emotion and motivation. En J. T., Cacciopo, L. G. Tassinary y G. G. Berntson (Eds.), Handbook of Psychophysiology (pp. 602-642). Nueva York: Cambridge University.
Brown, I. S., Kalish, H. I. y Farber, I. E. (1951). Conditioned fear as revealed by magnitude of startle response to an auditory stimulus. Journal of Experimental Psychology, 32, 317-328.
Cannon, W. B. (1931). Again the James-Lange an the talamic theories of emotion. Psychological Review, 38, 281-295.
Cobos, P., Sánchez, M., García, C., Vera, M. N. y Vila, J. (2002). Revisiting the James versus Cannon emotion: startle and autonomic modulatiion in patients with spinal cord injuries. Biological Psychology, 61, 251-269.
Dana, C. L. (1921). The anatomic seat of the emotions: a discussion of the James-Lange theory. Archives of Neurology and Psychiatry, 6, 634-639.
Davis, M. (1992). The role of the amygdala in conditioned fear. En J. P. Aggleton (Ed.), The amigdala: Neurobiological Aspects of Emotion, Memory, and Mental Disfunction. Nueva York: Wiley-Liss.
Davis, M. (1997). The neurobiological basis of acoustic startle modulation: Research on fear motivation. En P. J. Lang, R. F. Simons y M. Balaban (Eds.) Attention and orienting: Sensory and motivational processes (pp. 97-136).Londres: Lawrence Erlbaum.
Eysenck, H. J. (1960). The structure of human personality. Londres: Methuen.
Fernández, M. C., y Vila, J. (1989). Sympathetic-parasympathetic mediation of the cardiac defense response in humans. Biological Psychology, 28, 123-133.
Hohmann, G. W. (1966). Some effects of spinal cord lesion on experienced emotional feelings. Psychophysiology, 3, 526-534.
James, W. (1884). What is an emotion? Mind, 9, 188-205. (Traducción: Qué es una emoción?. Estudios de Psicología, 1985, nº 21).
Landis, C., y Hunt, W. (1939). The startle pattern. Nueva York: Farrar.
Lang, P. J. (1979). A bio-informational theory of emotional imagery. Psychophysiology, 16, 495-512.
Lang, P. J. (1995). The emotion probe: studies of motivation and attention. American Psychologist, 50, 372-385.
Lang, P. J., Davis, M., y Öhman, A. (2002). Fear and anxiety: Animal models and human cognitive psychophysiology. En L. Bäckman y C. von Hofsten (Eds.) Psychology at the turn of the millennium (vol. 1, pp. 377-411). Hove: Taylor & Francis.
LeDoux, J. E. (1996). The Emotional Brain: The mysterious underpinnings of emotional life. Nueva York: Simons and Shuster. (Traducción al castellano, 1999, Madrid: Planeta).
LeDoux, J. E. (2003). Synaptic self: How our brains become who we are. London: Penguin Books.
Mineka, S. (1992). Evolutionary memories, emotional processing, and emotional disorders. The Psychology of Learning and Motivation, 28, 161-206.
Moltó, J., Montañés, S., Poy, R., Segarra, P., Pastor, M. C., Tormo, M. P., Ramírez, I., Hernández, M. A., Sánchez, M., Fernández, M. C., & Vila, J. (1999). Un nuevo método para el estudio experimental de las emociones. El International Affective Picture System (IAPS). Adaptación española. Revista de Psicología General y Aplicada, 52, 55-87.
Morris, J. S., Öhman, A., y Dolan, R. J. (1998). Conscious and unconscious emotional learning in the human amygdala. Nature, 393, 467-470.
Newman, E. B., Perkins, F. T., y Wheeler, R. H. (1930). Cannon’s theory of emotion: A critique. Psychological Review, 37, 305-326.
Öhman, A., y Mineka, S. (2001). Fears, phobias, and preparedness: Towaqrd an evolved module of fear and fear learning. Psychological Review, 108, 483-522.
Osgood, C. E., Suci, G., y Tannenbaum, P. (1957). The measurement of meaning. En J. G. Snider y C. E. Osgood (Eds.) Semantic differential technique (pp. 3-41). Chicago, IL: Aldine.
Posner, M. I. (1999). Educación y Cerebro Humano. Discurso investidura Doctor "Honoris Causa". Granada: Servicio de Publicaciones.
Reyes del Paso, G. A., Godoy, J., y Vila, J. (1993). Respiratory sinus arrhytmia as an index of parasympathetic cardiac control during the cardiac defense response. Biological Psychology, 35, 17-35.
Reyes del Paso, G. A., Vila, J., y García, A. (1994). Physiological significance of the defense response to intense auditory stimulation. A pharmacological blockade study. International Journal of Psychophysiology, 15, 15-26.
Ruiz-Padial, E., Sánchez, M. B., Thayer, J. F., y Vila, J. (2002). Modulación no consciente de la respuesta cardíaca de defensa por imágenes fóbicas. Psicothema, 14, 739-745.
Ruiz-Padial, E. (2003). Modulación preatencional de reflejos defensivos. Tesis Doctoral. Universidad de Granada.
Russell, J. (1980). A circumplex model of affect. Journal of Personality and Social Psychology, 39, 1161-1178.
Sánchez, M., Ruiz-Padial, E., Pérez, N., Fernández, M. C., Cobos, P., & Vila, J. (2002). Modulación emocional de los reflejos defensivos mediante visualización de imágenes afectivas. Psicothema, 14, 702-707.
Thayer, J. F., y Siegle, G. J. (2002). Neurovisceral integration in cardiac and emotional regulation. IEE Engineering in Medicine and Biology, 21, 24-29.
Thurstone, L. L. (1931). The measurement of attitudes. Jounal of Abnormal Psychology, 26, 249-269.
Vila, J., Fernández, M. C., Pegalajar, J., Vera, M. N., Robles, H., Pérez, N., Sánchez, M. B., Ramírez, I., y Ruiz-Padial, E. (2003). A New Look at Cardiac Defense: Attention or Emotion?. The Spanish Journal of Psychology, 6,, 60-78.
Vila, J., Sánchez, M., Ramírez, I., Fernández, M. C., Cobos, P., Rodríguez, S., Muñoz, M. A., Tormo, M. P., Herrero, M., Segarra, P., Pastor, M. C., Montañés, S., Poy, R., y Moltó, J. (2001). El sistema internacional de imágenes afectivas (IAPS): Adaptación española. Segunda parte. Revista de Psicología General y Aplicada, 54, 635-657.
Whalen, P. J., Rauch, S. L., Etcoff, N. L., McInerney, S. C., Lee, M. B., y Jenike, M. A. (1998). Masked presentations of emotional facial expressions modulate amygdala activity withour explicit knowlege. The Journal of Neuroscience, 18, 411-418.
Vrana, S. R., Spence, F. L., y Lang, P. J. (1988). The startle probe response: A new measure of emotion? Journal of Abnormal Psychology, 97, 487-491.
[1] Este trabajo reproduce partes del capítulo 2 del libro Tratamientos Psicológicos: La aproximación experimental de Jaime Vila y María del Carmen Fernández-Santaella (en prensa).
[2] Las investigaciones que se mencionan en este trabajo han sido financiadas por el Ministerio de Ciencia y Tecnología (Proyectos PB97-0841 y BSO2001-3211) y la Junta de Andalucía (Grupo HUM-388).
|
|
Volver a la Reme |